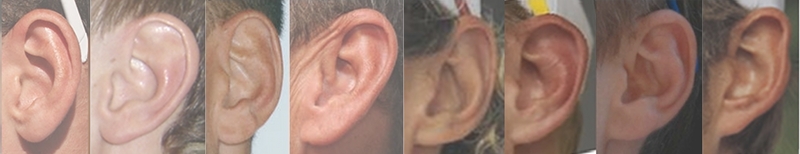
Earwax type: The myth
Some people have earwax that is wet, sticky and yellow or brown; other people's earwax is dry, crumbly and grayish. Variation at a single gene determines which kind of earwax you have; the allele for wet earwax is dominant over the allele for dry earwax. The allele for dry earwax appears to have originated by mutation in northeastern Asia about 2,000 generations ago, then spread outwards because it was favored by natural selection. It is very common in eastern Asia, becomes much less common towards Europe, and is very rare in Africa.
Earwax type is not used very often to illustrate basic genetics, but unlike most human characters that are used (tongue rolling, attached earlobes, etc.), it really is controlled by a single gene with two alleles.
The reality
Earwax as a character

Earwax, also known as "cerumen," is produced by glands that line the ear canal. It gradually moves out of the ear, carrying dead skin cells and other debris. Most people have one of two types of earwax. Wet earwax is sticky and yellowish brown to dark brown in color, while dry earwax is crumbly and gray to tan. A small number of people, generally less than 1 percent, have earwax that is intermediate or unclassifiable (Matsunaga 1962, Petrakis et al. 1967; Ing et al. 1973; Petrakis et al. 1990). Petrakis et al. (1986) found that the proportion of white women with dry earwax increased with age, which would suggest that earwax type was not determined solely by genetics, but Nakajima and Hirano (1969) did not find any change with age in Japan.
Family studies
Matsunaga (1962) wrote the first English-language paper on the polymorphism in earwax type, reviewing a large body of Japanese-language literature that dated back to the early 1930s. Data on Japanese families showed the following, where "W" is wet earwax and "D" is dry earwax:
| Parents | W offspring | D offspring |
|---|---|---|
| W x W | 35 | 12 |
| W x D | 205 | 195 |
| D x D | 0 | 634 |
If the allele for dry is recessive, two parents with dry earwax could not have a child with wet earwax, and this is what the data show. In addition, because the allele for wet earwax is rare in this sample, most W parents would be WD heterozygotes. You would then expect about a 3:1 ratio of W to D in offspring of WxW matings, and about a 1:1 ratio of W to D in WxD matings. The data also fit this prediction. These data support the conclusion that earwax is one-locus, two-allele trait, with the W allele being dominant.
Petrakis et al. (1967) performed a similar study in American Indians, with similar results:
| Parents | W offspring | D offspring |
|---|---|---|
| W x W | 32 | 6 |
| W x D | 20 | 9 |
| D x D | 0 | 42 |
and so did Nakajima and Hirano (1969) in Japan:
| Parents | W offspring | D offspring |
|---|---|---|
| W x W | 27 | 3 |
| W x D | 137 | 109 |
| D x D | 0 | 345 |
Molecular genetics
Tomita et al. (2002) used eight Japanese families to determine that the gene for wet/dry earwax is on chromosome 16, near the centromere. Yoshiura et al. (2006) then found the gene responsible: ABCC11 (ATP-binding cassette, subfamily C, member 11). The allele for wet earwax has a G at site 538 of the coding region, which causes a glycine at position 180 in the amino acid sequence; most dry alleles have an A at site 538, coding for arginine. There is another, less common allele, a deletion of 27 nucleotides in exon 29, that also causes dry earwax. Toyoda et al. (2009) stained the ABCC11 protein in various tissues and found that there was less of the protein in individuals with the AA (dry earwax) genotype, suggesting that the A allele causes a loss of function of the ABCC11 protein.
Ile Ala Ser Val Leu Gly Pro Ile Leu Ile Ile Pro
wet earwax ATT GCC AGT GTA CTC GGG CCA ATA TTG ATT ATA CCA
dry earwax ATT GCC AGT GTA CTC AGG CCA ATA TTG ATT ATA CCA
Arg
Bases 523-558 of the coding sequence of ATCC11, along with the amino acid sequence. The DNA polymorphism at site 538 causes the amino acid polymorphism that determines earwax type.
The allele for dry earwax has frequencies close to 100% in people from northern China and Korea, is intermediate in people from Japan, southern Asia, and the Americas, is uncommon in Europe, and is almost completely absent in Africa (Petrakis et al. 1971, Ibraimov 1991, Yoshiura et al. 2006, Ohashi et al. 2011). There are very few polymorphisms in the human genome that show this much difference in allele frequency between one geographic area and another. Ohashi et al. (2011) used patterns of variation at two nearby microsatellite loci and estimated that the allele for dry earwax originated as a new mutation about 2000 generations ago and has spread due to natural selection, with dry earwax individuals having a relative fitness about 1 percent higher than wet earwax individuals. Wet earwax is associated with warmer areas in Europe, Asia and the Americas (Ohashi et al. 2011), so there seems to be something about cooler climates that favors the dry earwax allele.
Earwax and body odor
There are two kinds of sweat glands: eccrine sweat glands, which are found throughout the skin, and apocrine sweat glands, which are found in the armpits and groin. Eccrine sweat glands produce sweat that is mostly water and salt, and it does not contribute very much to body odor. Apocrine sweat contains proteins and lipids; when bacteria on the skin metabolize apocrine sweat, they produce body odor. The earwax glands (ceruminous glands) are a form of apocrine gland.
In Japan, wet earwax has long been associated with greater body odor (Adachi 1937, Matsunaga 1962). Yoo et al. (2006) found that 860 out of 896 patients who consulted a doctor about axillary osmidrosis (the medical term for stinky armpits) had wet earwax, in a population where wet earwax is uncommon. Nakano et al. (2009) genotyped the polymorphism in the ABCC11 gene in Japanese axillary osmidrosis patients and found the genotypes that cause wet earwax in 78 out of 79, while only 35 percent of the general population had wet earwax genotypes; Inone et al. (2010) found similar results in a smaller study. Martin et al (2010) analyzed sweat from people of different ABCC11 genotypes and found that several compounds that are precursors for body odor are absent or in lower concentrations in sweat from people with the dry earwax genotype.
Earwax and breast cancer
Like the earwax glands, the mammary glands are a form of apocrine gland, and the ABCC11 gene is expressed in both. Petrakis (1971) noticed that breast cancer was less common in geographic areas with high frequencies of dry earwax. He found a higher frequency of wet earwax in Japanese-American women with breast cancer than in a control group of Japanese-American women, although the difference was not statistically significant. Ing et al. (1973) surveyed a much larger sample of breast cancer patients and controls in Hong Kong and did not find a significant difference in the frequency of wet earwax.
Ota et al. (2010) compared ABCC11 genotypes in Japanese breast cancer patients and controls and did find a significantly higher frequency of the allele for wet earwax in breast cancer patients; the genotypes for wet earwax were in 25 percent of the cancer patients and 17 percent of the controls. There was no difference in ABCC11 genotype frequency between breast cancer patients and controls in large samples of German (Lang et al. 2011), Australian or Italian (Beesley et al. 2011) women, but fewer than 2 percent of women in those populations have dry earwax.
There is additional evidence that the ABCC11 polymorphism affects the mammary glands. Breast fluids can be collected from some women who are not producing milk, and Petrakis et al. (1981) found that both Asian and Caucasian women with wet earwax were more likely to produce breast fluids. Petrakis et al. (1990) confirmed this pattern, but found that there was a larger difference between wet and dry earwax women for Asian immigrants to the United States than for U.S.-born Asian-Americans, suggesting that the environment also plays a role. Miura et al. (2007) found that women with dry earwax were less likely to produce colostrum (the breast milk produced around childbirth), and those who did produced smaller volumes than women with wet earwax.
Conclusion
Unlike most of the human characters that are used to demonstrate simple genetics principles, wet vs. dry earwax really is controlled by one gene with two alleles. Several factors make it especially attractive as a classroom topic: the gene has been identified, the biochemical basis for the variation is fairly well understood, there is evidence for strong natural selection, and there are links to human health (body odor and possibly cancer). There are, however, two problems with using earwax type in classroom exercises. One is that the allele for dry earwax has a frequency in western European populations of 10 to 20 percent, and it is virtually absent from African populations. Because dry earwax is recessive, this means that a classroom of students of European and African ancestry may show no variation; everyone could have wet earwax. Classrooms in northern China or Korea, on the other hand, may contain only students with dry earwax, because the allele for dry earwax is at nearly 100 percent in those areas.
The second problem with using earwax type in a genetics exercise is that having a bunch of students sitting in a classroom, digging wax out of their ears and waving it around, would be kind of disgusting.
References
Adachi, B. 1937. Das Ohrenschmalz als Rassenmerkmal und der Rassengeruch ("Achselgeruch") nebst dem Rassenunterschied der Schweissdrüsen. Zeitschrift für Rassenkunde und die Gesamte Forschung am Menschen 6: 273-307.
Beesley, J., S. E. Johnatty, X. Q. Chen, A. B. Spurdle, P. Peterlongo, M. Barile, V. Pensotti, S. Manoukian, P. Radice, and G. Chenevix-Trench. 2011. No evidence for an association between the earwax-associated polymorphism in ABCC11 and breast cancer risk in Caucasian women. Breast Cancer Research and Treatment 126: 235-239.
Ibraimov, A.I. 1991. Cerumen phenotypes in certain populations of Eurasia and Africa. American Journal of Physical Anthropology 84: 209-211.
Ing, R., L. Petrakis, and H. C. Ho. 1973. Evidence against association between wet cerumen and breast cancer. Lancet 1: 41.
Inone, Y., T. Mori, Y. Toyoda, A. Sakurai, T. Ishikawa, Y. Mitani, Y. Hayashizaki, Y. Yoshimura, H. Kurahashi, and Y. Sakai. 2010. Correlation of axillary osmidrosis to a SNP in the ABCC11 gene determined by the Smart Amplification Process (SmartAmp) method. Journal of Plastic, Reconstructive and Aesthetic Surgery 63: 1369-1374.
Lang, T., et al. (13 co-authors). 2011. The earwax-associated SNP c.538G>A (G180R) in ABCC11 is not associated with breast cancer risk in Europeans. Breast Cancer Research and Treatment, e-pub ahead of print.
Martin, A., M. Saathoff, F. Kuhn, H. Max, L. Terstegen, and A. Natsch. 2010. A functional ABCC11 allele is essential in the biochemical formation of human axillary odor. Journal of Investigative Dermatology 130: 529�540.
Matsunaga, E. 1962. The dimorphism in human normal cerumen. Annals of Human Genetics 25: 273-286.
Miura, K., K. Yoshiura, S. Miura, T. Shimada, K. Yamasaki, A. Yoshida, D. Nakayama, Y. Shibata, N. Niikawa, and H. Masuzaki. 2007. A strong association between human earwax-type and apocrine colostrum secretion from the mammary gland. Human Genetics 121: 631�633.
Nakajima, A., and I. Hirano. 1969. Distribution and inheritance of earwax types: a study on inhabitants in Awa District, Chiba Prefecture. Japanese Journal of Human Genetics 13: 201-207.
Nakano, M., N. Miwa, A. Hirano, K. Yoshiura, and N. Niikawa. 2009. A strong association of axillary osmidrosis with the wet earwax type determined by genotyping of the ABCC11 gene. BMC Genetics 10:42.
Ohashi, J., I. Naka, and N. Tsuchiya. 2011. The impact of natural selection on an ABCC11 SNP determining earwax type. Molecular Biology and Evolution 28: 849-857.
Ota, I. et al. (17 co-authors). 2010. Association between breast cancer risk and the wild-type allele of human ABC transporter ABCC11. Anticancer Research 30: 5189-5194.
Petrakis, N.L. 1971. Cerumen genetics and human breast cancer. Science 173: 347-349.
Petrakis, N. L., K. T. Molohon, and D. J. Tepper. 1967. Cerumen in American Indians: genetic implications of sticky and dry types. Science 158: 1192-1193.
Petrakis, N.L., U. Pingle, S. J. Petrakis, and S. L. Petrakis. 1971. Evidence for a genetic cline in earwax types in Middle East and south Asia. American Journal of Physical Anthropology 35: 141-144.
Petrakis, N. L., V. L. Ernster, S. T. Sacks, E. B. King, R. J. Schweitzer, T. K. Hunt, M.-C. King. 1981. Epidemiology of breast secretion: Association with breast cancer risk factors and cerumen type. Journal of the National Cancer Institute 67: 277-284.
Petrakis, N.L., S. L. Wiesenfeld, and L. Flander. 1986. Possible influence of age on the expression of the heterozygous cerumen phenotype. American Journal of Physical Anthropology 69: 437-440.
Petrakis, N. L., E. B. King, M. Lee, and R. Miike. 1990. Cerumen phenotype and proliferative epithelium in breast fluids of United States-born vs. immigrant Asian women: a possible genetic-environmental interaction. Breast Cancer Research and Treatment 16: 279-285.
Tomita, H., K. Yamada, M. Ghadami, T. Ogura, Y. Yanai, K. Nakatomi, M. Sadamatsu, A. Masui, N. Kato, and N. Niikawa. 2002. Mapping of the wet/dry earwax locus to the pericentromeric region of chromosome 16. Lancet 359: 2000-2002.
Toyoda, Y., et al. (12 co-authors). 2009. Earwax, osmidrosis, and breast cancer: why does one SNP (538 g> a) in the human ABC transporter ABCC11 gene determine earwax type? FASEB Journal 23:2001�2013.
Yoo, W. M., N. S. Pae, S. J. Lee, T. S. Roh, S. Chung, and K. C. Tark. 2006. Endoscopy-assisted ultrasonic surgical aspiration of axillary osmidrosis: a retrospective review of 896 consecutive patients from 1998 to 2004. Journal of Plastic, Reconstructive and Aesthetic Surgery 59: 978-982.
Yoshiura, K., et al. (39 co-authors). 2006. A SNP in the ABCC11 gene is the determinant of human earwax type. Nature Genetics 38: 324-330.
OMIM entry on earwax and OMIM entry on ABCC11
Return to John McDonald's home page
This page was last revised September 16, 2013. Its address is http://udel.edu/~mcdonald/mythearwax.html. It may be cited as pp. 28-33 in: McDonald, J.H. 2011. Myths of Human Genetics. Sparky House Publishing, Baltimore, Maryland.
©2011 by John H. McDonald. You can probably do what you want with this content; see the permissions page for details.