Current Research Projects
1. Models of "Vulnerable" Atherosclerotic Plaque
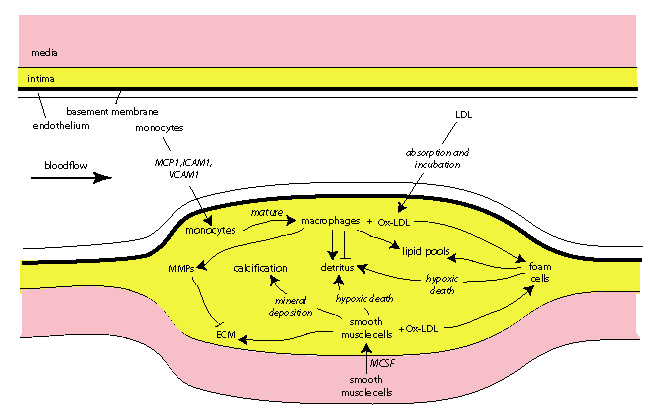
Cardiovascular disease (CVD)
affected about 80 million Americans in 2006 and was responsible for
800,000 deaths. One common form of CVD is atherosclerosis which is a ``furring'' of the artery
walls leading to a narrowing of the vessel lumen. It was originally
thought that plaque build-up would make patients more
susceptible to Myocardial Infarction (MI) or heart attacks. However, doctors
now think that the rupture of certain plaques called
Thin Capped Fibroatheromas or ''vulnerable plaques'' are responsible for most
fatalities. In one study, 73% of all deaths from MI were
caused by plaque rupture. This has led to the definition and characterization of
vulnerable plaque which is thought to be the
major culprit behind MIs.
The formation of plaque is a very complicated process. Some of the important
factors are shown in the figure above. Our group is
trying to quantify plaque growth in order to aid in the detection and diagnosis
of plaque. Our research aims to answer some
important basic questions concerning plaque growth. For example, how
do shear stresses affect the leukocyte adhesion cascade (LAC) and, subsequently,
how does the LAC affect plaque formation? Which factors determine
whether a plaque remodels inwardly or outwardly? Why do necrotic
cores form inside vulnerable plaques and what determines their shape?
Some relevant literature
- The Vulnerable Atherosclerotic Plaque: Strategies for diagnosis and management,
edited by R. Virmani, J. Narula, M. B. Leon, and J. T. Willerson. Blackwell 2007.
- Shah, P. K., Mechanisms of Plaque Vulnerability and Rupture, Journal of the American
College of Cardiology 41, No. 4 (2003).
- Virmani et al., Lessons from sudden coronary death: A comprehensive morphological classification
scheme for atherosclerotic lesions. Arteriosclerosis, Thrombosis and Vascular Biology 20
pp 1262-1275 (2000).
- T. I. Zohdi, G. A. Holzapfel and S. A. Berger, A phenomenological model for atherosclerotic
plaque growth and rupture. Journal of Theoretical Biology 227 pp 437-443 (2004).
Here are some movies generated by computer simulation, courtesy of Mr Navid Mohammad Mirzaei, my PhD student:
Movie A
Movie B
Movie C
Movie D
Movie E
2. Stochastic Inverse Problems
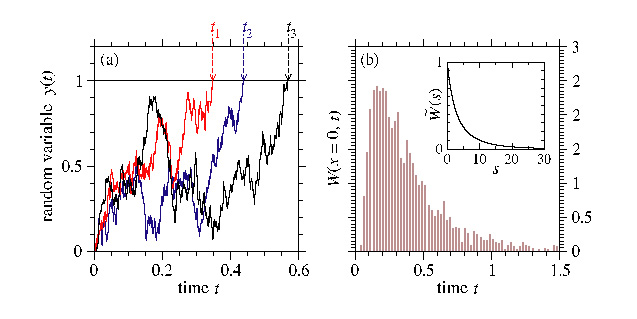
The study of stochastic processes is important
in many branches of mathematics, science
and engineering. One example of a stochastic process
is a (persistant) brownian motion where changes in a walker's position
arise from two components: a deterministic drift and a random noise.
Now consider a situation where one only
has knowledge of the exit times of a random walk, i.e. one
only knows the times at which the random walker reaches a certain
value. Is it possible to infer the drift from these times?
This problem is a simple example of a Stochastic Inverse Problem .
Such problems have great importance in biology.
For example, under certain modeling assumptions, they can be used to
interpret Dynamic Force Spectroscopy (DFS) data
in the context of protein folding and chemical
bond rupture.
In DFS experiments, an Atomic Force Microscope is used to
slowly unfold a macromolecule. Because thermal fluctuations
are important at such small scales, the data generated by such experiments consists
not of a single rupture force, but a distribution of such forces. One active
area of research is to interpret these data sets. What can the distribution
of rupture forces tell us about how macromolecules fold and unfold?
The extent to which a macromolecule has unfolded can be described in terms
its bond coordinate which, qualitatively, can be thought of as a measure
of its end-to-end distance. Often, the unfolding can be modeled as a random
walk for its bond coordinate. A DFS experiment can be thought of
as an activated escape of a brownian particle from a potential well, which
accounts for all the
intramolecular forces that hold the molecule in a
particular configuration. The goal then
is to infer the shape of the potential well from rupture time/force data.
Some relevant literature:
- P.-W. Fok and T. Chou, Reconstruction of potential energy profiles from multiple rupture
time distributions, Proceedings of the Royal Society A 466, No. 2124 pp 3479-3499 (2010)
- Bal, G. and Chou, T., On the reconstruction of diffusions
using a single first-exit time distribution. Inverse Problems
20, 1053-1065 (2003).
- Dudko, O. K., Hummer, G. and Szabo, A., Theory, analysis,
and interpretation of single-molecule force spectroscopy experiments.
Proc Natl Acad Sci USA 105, 15755-15760 (2008).
- Evans, E., Ritchie, K. and Merkel, R. Sensitive force technique to probe molecular adhesion and
structural linkages at biological interfaces. Biophysical Journal 68, 2580-2587 (1995).
- Freund, L. B. Characterizing the resistance
generated by a molecular bond as it is forcibly separated. Proc
Natl Acad Sci USA 106, 8818-8823 (2009).
Previous Research Projects
3. DNA Search and Repair
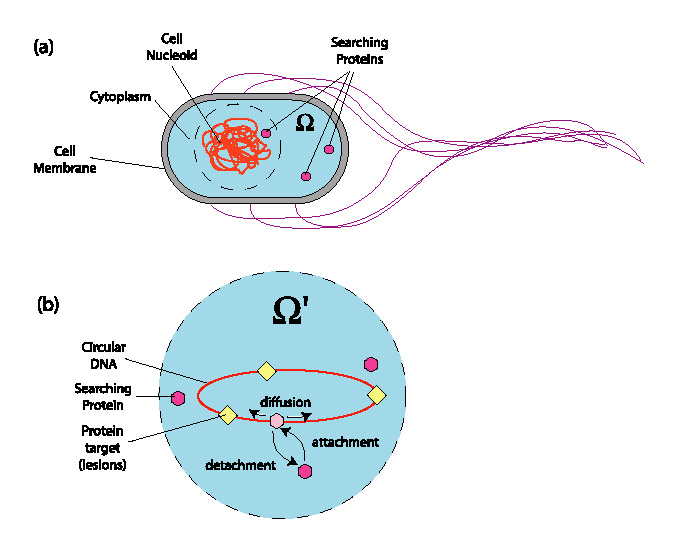
DNA repair enzymes are responsible for locating and
repairing damaged bases on DNA. Without them, mutations
in the DNA can occur, ultimately giving rise to diseases
like cancer. An important problem that has puzzled biophysicists
since the 70s is how enzymes are able to find small targets
(such as damaged bases) on a long DNA strand so rapidly.
For example the time to perform a 1D random search along a DNA strand,
with the enzyme permanently associated with the DNA, is
so long that it exceeds even the life cycle of the cell!
Likewise, a 3D random search within the
volume of a cell nucleus (or within the cell itself for prokaryotes)
yields search times that are inconsistent with experimental
reaction rates.
In the 1980s, a theory called facilitated diffusion was proposed
to explain fast search times for searching enzymes. The theory
posited that up to a 100 fold speed-up in the search could occur
if a combination of 1D and 3D searches was used. However
facilitated diffusion requires certain conditions to be met.
For example, the enzyme must spend half the search time in 1D and half
in 3D. These conditions are seldom met in practice. Subsequently
many researchers have tried to refine and extend
the theory to real biological systems.
Some relevant literature:
- P.-W. Fok and T. Chou, Accelerated search
kinetics mediated by redox reactions of DNA repair enzymes,
Biophysical Journal 96, pp 2949-3958 (2009).
- P.-W. Fok and T. Chou, Charge transport mediated
recruitment of DNA repair enzymes. J. Chem. Phys. 129 235101 (2008).
- O.G. Berg, R.B. Winter and P.H. Von Hippel,
Diffusion-driven mechanism of protein translocation
on nucleic acids. 1. Models and theory. Biochemistry 20
pp 6929-6948 (1981).
- M. Slutsky and L. A. Mirny, Kinetics of protein-DNA
interaction: facilitated target location in sequence-dependent
potential. Biophysical Journal 87 pp 4021 - 4035 (2004).
- T. A. Hu, A. Y. Grosberg and B. I. Shklovskii, How
proteins search for their specific sites on DNA: the role
of DNA conformation. Biophysical Journal 80 pp 2731-2744.
4. Epitaxial Growth and Step Models
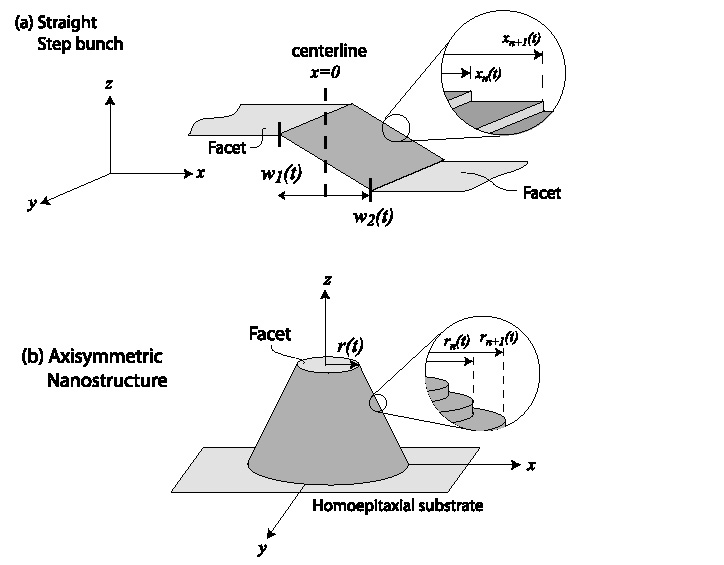
A vicinal surface is one that results
from a slight miscut from a high symmetry plane of a
crystal. As a result, the evolution of the surface is mediated
by the motion of discrete atomic-height steps. These steps
change their position in response to attachment-detachment
kinetics at step edges and diffusion on terraces (the region
between steps). For relaxing axisymmetric
nanostructures under near-equilibrium conditions, the circular
steps of these nanostructures are subject to two kinds of physical
effect. First, the radii of steps shrink because of step line tension.
Second, step-step repulsions (stemming from entropic and/or elastic
interactions) ensure that steps cannot
get too close to each other. As relaxation proceeds, a circular
facet develops on the top of the structure and expands as the
inner most steps repeatedly collapse.
It has been known for some
time (e.g. see publication #4 below) that
within a macroscopic treatment,
nanostructure relaxation below the roughening
temperature is described by
nonlinear PDEs that are singular at facet edges.
These singularities are intimately related to the
non-analyticity of the surface free energy below
the roughening temperature. One way to
to solve these problems is to regard the facet
edge as a free boundary whose position
is time dependent and unknown a priori (see publication #3).
The question that naturally arises is
what the boundary conditions should be at the facet edge.
In publication #1, we propose a boundary condition that
depends on the motion of the inner most step:
if the step collapse frequency
is large, the facet expands quickly. On the other hand, if the
inner most step does not move at all, the facet is frozen in time.
The boundary condition bridges two length scales,
coupling together the
downward macroscopic translation of the facet
with microscopic step collapses. Application of this boundary
condition yields
solutions that are in excellent agreement with
simulation results.
In publication #2, we study finite and semi-infinite collections of
straight and circular steps,
comparing the discrete equations to their respective continuum PDEs.
Boundary conditions
are derived from simple physical considerations at the facet edges.
For straight steps,
simple similarity solutions exist which -- in some sense -- provide an exact
description of the step motion.
The solutions have different scalings in time depending on whether
the system is semi-infinite or strictly finite.
For circular steps where step-step interactions are weak compared to step line tension,
the continuum PDE reduces to a kinematic wave equation.
Shockwave theories and the method of characteristics can be used to treat facet translation
in this case.
Some relevant literature
- D. Margetis, P.-W. Fok, M.J. Aziz and H.A. Stone, Continuum
theory of nanostructure decay via a microscale condition,
Physical Review Letters 97, 096102 (2006)
- P.-W. Fok, R. R. Rosales and D. Margetis,
Facet evolution of supported nanostructures: Effect of finite height
Physical Review B 78, 235401 (2008)
- H. Spohn, Surface dynamics below the roughening transition,
J. Phys. I France 3 pp 69-81 (1993)
- A. Rettori and J. Villain, Flattening of grooves on
a crystal surface: a method of investigation of surface
roughness. J. Phys. France 49 pp 257-267 (1988)
5. Multirate Integration Schemes
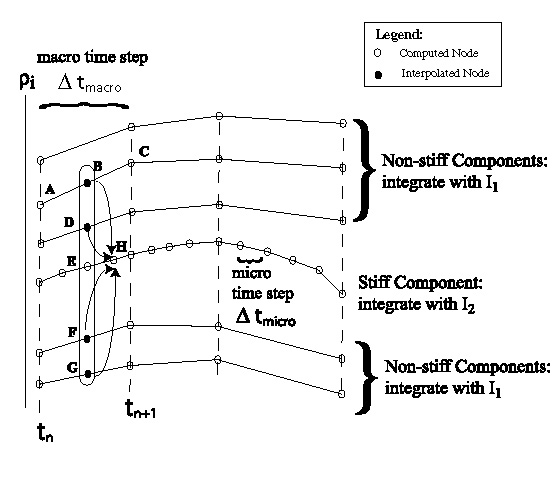
Multirate integration of a set of Ordinary Differential
Equations (ODEs) involves taking time steps of different size
for different components in the solution. This strategy
is especially useful when a small fraction of the components
evolves on faster time scales than the rest.
One of the main difficulties with implementing multirate
methods is that slower components in the ODE system
have to be interpolated accurately. Another difficulty
is that it is unclear what order of interpolant is needed
if the multirate method is to be of nth order.
Can this interpolation be implemented without using any
extra function evaluations from the ODE?
Our group is currently developing
a 4th order multirate method for locally coupled ODEs.
Our aim is to apply this
method to a wide variety of PDEs that have been discretized
using the method of lines.
Some relevant literature
- C.W. Gear and D.R. Wells, Multirate linear multistep methods,
BIT 24 484 (1984).
- A. Logg, Multi-adaptive time integration, Appl. Numer. Math 48
pp 339-354 (2004).
- V. Savcenco, W. Hundsdorfer and J.G. Verwer, A multirate time
stepping strategy for stiff ordinary differential equations, BIT 47
pp 137-155 (2007).
- W. Hundsdorfer and V. Savcenco, Analysis of a multirate
theta-method for stiff ODEs, Applied Numerical Mathematics 59
pp 693-706 (2008).
|